
Research
1. The mechanisms of Echinoid-mediated cell adhesion and migration
Members of the immunoglobulin (Ig) superfamily, cadherins and integrins represent the three major classical groups of cell adhesion molecules (CAMs) and play important roles in various developmental processes in all vertebrate and invertebrate species. Echinoid (Ed) is an immunoglobulin domain-containing CAMs and participates in either homophilic or heterophilic interactions. Ed negatively regulates the EGFR signaling pathway during Drosophila eye development and it facilitates Notch signaling during adult sensory bristle patterning. Recently, we found that Ed is a novel component of Adherens Junctions (AJs) where cadherins are the major Ca2+-dependent homophilic CAM to mediate cell-cell adhesion. By possessing a C-terminal PDZ domain-binding motif, Ed binds to and localizes cell polarity protein Bazooka (Baz)/PAR-3, and F-actin-binding protein Canoe (Cno)/AF-6/afadin to AJs. Cells lacking either Ed or DE-Cadherin (DE-cad) accumulate a high density of the reciprocal protein. Thus Ed cooperates with DE-Cad to mediate cell adhesion. We are studying the mechanism of Ed-mediated cell adhesion and cell migration in various developmental processes.
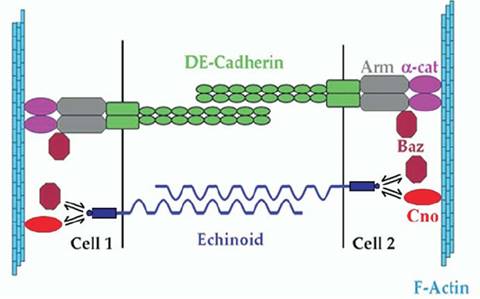
Model for Ed and DE-Cad sharing similar functions in cell-cell adhesion. Both Ed and DE-Cad form homophilic interactions between neighbouring cells. Ed uses its C-terminal EIIV motif to localize Baz and Cno to AJs. Cno, in turn, associates with F-Actin. This is similar to DE-Cad, which uses Arm and a -catenin to bind Baz and F-Actin, respectively.
2. Weckle is a zinc finger adaptor of the Toll pathway in dorso/ventral patterning of the Drosophila embryo
The Drosophila Toll pathway takes part in both establishment of the embryonic dorsoventral axis and induction of the innate immune response in adults. Upon activation by the cytokine Spatzle, Toll interacts with the adaptor proteins DmMyD88 and Tube and the kinase Pelle, and triggers degradation of the inhibitor Cactus thus allowing the nuclear translocation of the transcription factor Dorsal/Dif. weckle (wek) was previously identified as a new dorsal group gene that encodes a putative zinc-finger transcription factor. We demonstrated that cactus is epistatic to wek that in turn is epistatic to Toll . Consistent with this, Wek localizes to the plasma membrane of embryos, independent of Toll signaling. Wek homodimerizes and associates with Toll. Moreover, Wek binds to and localizes DmMyD88 to the plasma membrane. Thus, Wek acts as an adaptor to link Toll and DmMyD88 and therefore, to assemble/stabilize a Toll/Wek /DmMyD88/Tube complex. Remarkably, unlike the DmMyD88/tube/pelle/cactus gene cassette of the Toll pathway, wek play minimal role, if any, in the immune defense against Gram-positive bacteria and fungi. These results reveal differences in the Toll-mediated activation of Dorsal in the embryo and Dif in the fat body of adult flies.
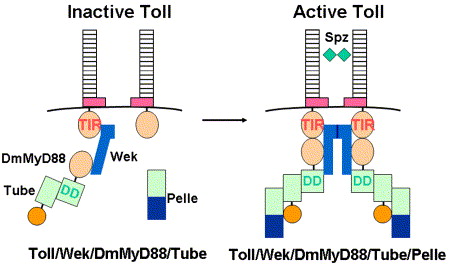
Model for molecular interactions between components of the Toll pathway. Prior to Toll activation, Wek interacts with Toll and recruits DmMyD88/Tube complex to form a relatively unstable Toll/Wek/DmMyD88/Tube presignaling complex on the membrane. The cysteine-containing motif (pink) of Toll self-inhibits Toll activation. Upon Toll dimerization by Spatzle, active Toll simultaneously recruits both DmMyD88 and Wek two adaptors to form a more stable Toll/Wek/DmMyD88/Tube complex that activates Pelle. Wek can self-associate through WekN domain.