
蛋白質的故事:
螢火蟲螢光素酵素(firefly luciferase)
|
蛋白質的故事: 螢火蟲螢光素酵素(firefly luciferase) |
夏夜中的螢火蟲,尾部會發出一閃一閃的亮光,相當特別而引
人注目。螢火蟲為什麼會發光?其實不只是螢火蟲,許多其他的生物也會發出類似的螢光,統稱生物螢光(bioluminescence)。研究顯示這些螢光都是利用同一類的發光原理:一種蟲螢光素(luciferrin)和一種蟲螢光酵素(luciferase)的氧化還原反應。

蟲螢光素酵素在常溫、常壓下使發光反應進行非常有效率,和三磷酸腺甘(ATP)作用產生複雜的氧化還原反應,這種反應是連續性的進行著。螢火蟲的光沒有伴隨熱,能量和效率非常高。約2∼10 %的能量轉為熱量,而其餘能量完全用來發光,因此螢火蟲的光並不像電燈泡般燙人,故稱為「冷光」。
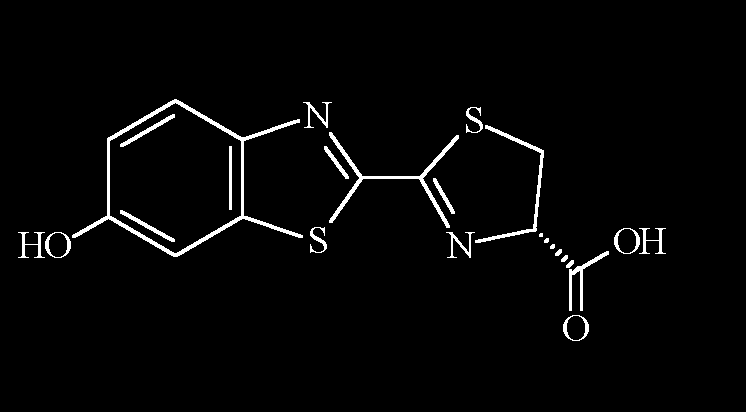
生物螢光中所使用的蟲螢光素和蟲螢光素酵素,目前已發現許多種不同的組合。螢火蟲所使用的蟲螢光素的結構式為:
螢火蟲的蟲螢光素酵素是一個分子量約 62 kDa 的氧化酵素(oxygenase) 但是和其他氧化酵素所不同的是,沒有額外的氧化還原互補官能基(redox prosthetic group)參與反應。螢火蟲的蟲螢光素酵素,其結構折疊成兩個不同的區域(domain)。結構中主要的部份 ,由殘基(residue)4 - 436 所組成,包含一個緊密的區域,其中又含有一個扭曲的反平行 β-桶形(antiparallel β-barrel),以及兩個β-平板(β-sheets),並有α-螺旋在旁側。此蛋白質的碳端(C-terminus),殘基 440 - 544,另外形成一個小α+β區域。
較大的氮端(N-terminal domain),兩個β-平板的子區域(subdomains)組合成五層 αβαβα的三級結構(tertiary structure)所以兩個α-螺旋像三明治一般被夾在平板之中,而其他的螺旋被包在相對於外邊的那一面。在這個酵素的結構中,兩個β-平板被弄傾斜,使得平行股(strand)的碳端可以互相靠近。 每個平板由八股組成,並顯示出向右扭轉的特性。每個平板的核心組成,由許多平行股和α-螺旋形成標準的右手交錯型連結(standard right-handed cross-over connections),結果就是螺旋重新排列在平板的兩側,反平行於各股。
碳端區域包含兩個短的反平行反平行 β-股(short antiparallel β-strands )以及一個三股混合的β-平板,三個螺旋包在對邊。這種折疊的類型已經被分類為一種α+β架構。
螢火蟲的蟲螢光素酵素的活性部位(active site)的可能位置,可以由三個相關酵素家族:螢火蟲的蟲螢光素酵素、醯基輔酵素A連接酵素(acyl-CoA ligase)、蛋白鏈合成酵素(peptide synthetase)的高度保存不變(highly conserved)的殘基位置來定出。