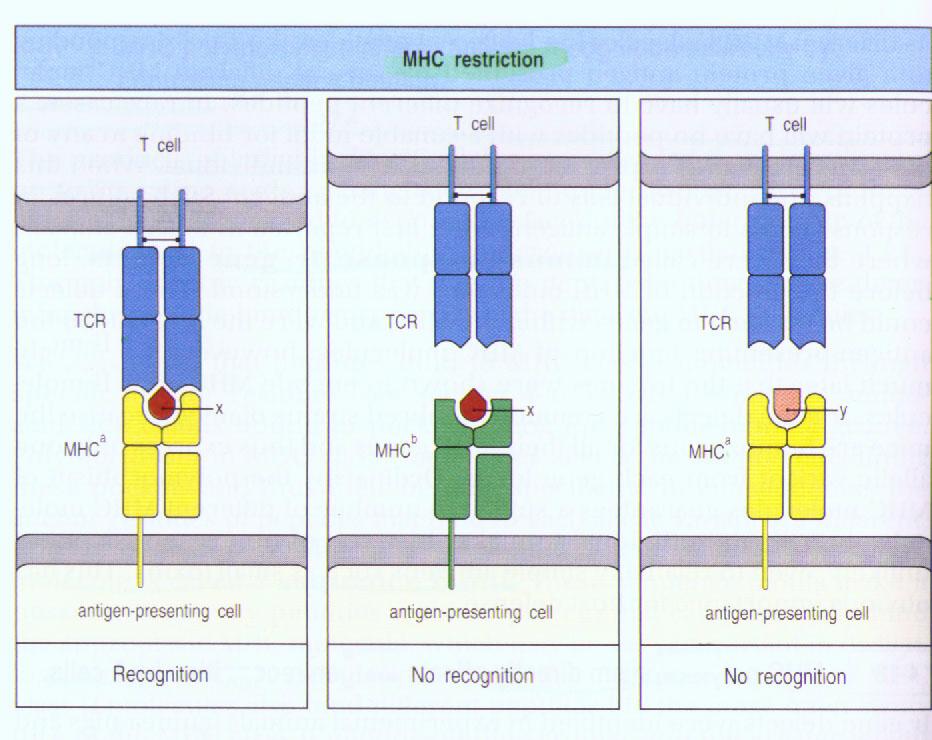
Ⅰ.αβT cell recognition of antigen is MHC-restricted.
Crystal Structure of a γδT cell Receptor Ligand T22:A Truncated MHC-Like Fold
Author: Chister Wingren, Michael P. Crowley, Massimo
Degano, Yueh-hsiu Chien, Lan
A.Wilson.
Source: Science, Vol 287, 14 January 2000
Speaker: 生科01 陳君柏 861610
Abstract:
Murine T10 and T22 are highly related nonclassical major histocompatibility complex (MHC)
class Ib proteins that bind to certain γδT cell receptors (TCRs) in the absence of other
components. The crystal structure of T22b at 3.1 angstroms reveals similarities to MHC
class I molecules, but one side of the normal peptide-binding groove is severely
truncated, which allows direct access to the -sheet floor. Potential TCR-binding sites can
be inferred from functional mapping of T10 and T22 point mutants and allelic variants.
Thus, T22 represents an unusual variant of the MHC-like fold and indicates that and TCRs
interact differently with their respective MHC ligands.
Keyword:
Ⅰ.αβT cell recognition of antigen is MHC-restricted.
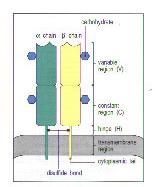
Ⅱ.αβ T cell receptor consists of two different polypeptide chains, termed the T-cell receptor α and β chains.
Ⅰ.γδT cells diverge from a common precursor identical to αβT cells.
Ⅱ.γδT cell receptor consists of γδ chains.
MHC(Major Histocompatibility Complex) Class 1a:
Ⅰ.MHC Class 1a is a heterodimer of a membrane-spanning αchain, non-covalently associated with β2-microglobulin, which dose not span the membrane.
Ⅱ.The most remarkable feature of MHC Class 1a molecules is the structure of the α1 and α2 domains, which pair to generate a craft on the surface of the molecular that is the site of peptide binding.
Ⅰ.MHC Class 1b molecules are β2-microglobulin-associated cell-surface molecule.
Ⅱ.The number of MHC Class 1b molecule is about 150 or more in human.
Ⅲ.Some MHC Class 1b molecules have specialized roles in antigen presentation and some may play a part in innate immunity or in the induction of immune responses.
Ⅰ.T22 is one of MHC Class 1b molecules and have been identified as specific ligand for two γδT cell clones, G8 and KN6.
Ⅱ.Recognition of T22 by G8 γδT cells is fundamentally different from MHC Class 1a recognition by αβT cells.
Ⅲ.T22 do not appear to require peptide or any other ligand for cell surface expression or recognition by γδT cells.
Ⅳ.Recognition and stimulation are dependent on a properly folded and stable heterodimer association of the T22 heavy chain with β2-microglobulin.
The comparison of three-dimensional structure between T22 and MHC class 1a molecule:
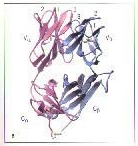
1.Both αchains fold into the three domains that are associated with β2M. A and C are T22; B and D are MHC class 1a.
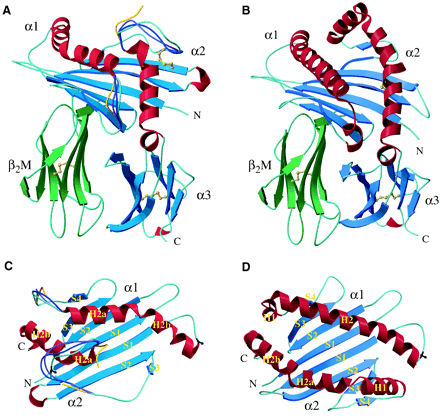
3.In MHC class 1a, the α1 domain has two α-helical segments, named H1 and H2. However, T22 lacks the entire H1 segment and does not start the H2 helical structure until residue 60
4.T22 lack the S4 βstrand and H1 helical segment in the α2 domain of MHC class 1a
5.The conformation of the hairpin loop connecting S1 and S2 betwwn S3 and H2
6.The distance between the α1 and α2 helices in T22 is shorter than that in MHC class 1a.
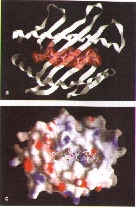
7.Analysis of the electrostatic surface potential reveals the top of the α1α2 domains to be mainly neutral. Positive potential is colored blue, neutral potential is colored gray, and negative potential is colored red(A). Potential γδT cell-binding sites on the α1α2 domains of T22 are showed (B). Two potential interaction sites for the γδ TCR are highlighted on the exposed β-sheet floor and on the acidic patch at the opposite edge of teh T22 surface.
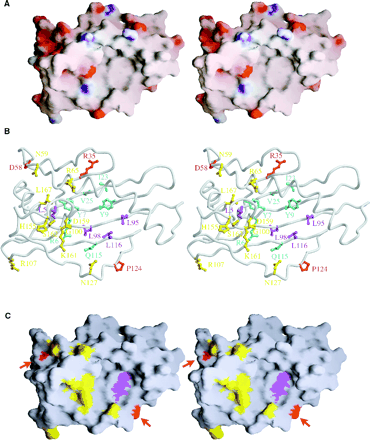
8.T22 has two probable N-linked glycosylation sites at residues 86 and 150 in the α1α2 domains
1.The structural differences between T22 and MHC class 1a molecules support the idea that γδ and αβ TCRs recognize antigens differntly.
2.γδ TCRs are not restricted in their choice of ligands or epitopes and independent of bound peptide and distinct from that recignized by peptide-specific αβ T cells.
3.The crystal struture of T22 reveals another way for the versatile MHC fold to adapt to a different function in the immune system.