以下為原始文件,由於擔心網路傳輸速度不佳以及資料的變動因子等等,所以在此收錄一份在 Nature 雜誌同樣的文件,以供有興趣了解第一手資料的您觀看。
順便告訴你一個祕密,在雜誌上的原文,附圖一的細胞圖片有錯誤, Nature 網路版的則有修正哦!
Nature, Volume 385, 810 - 813, February 27, 1997
I. Wilmut, A. E. Schnieke*, J. McWhir,
A. J. Kind* & K. H. S. Campbell
Roslin Institute (Edinburgh), Roslin,
Midlothian EH25 9PS, UK * PPL Therapeutics, Roslin,
Midlothian EH25 9PP, UK
It has long been known that in amphibians, nuclei transferred from adult keratinocytes established in culture support development to the juvenile, tadpole stage3. Although this involves differentiation into complex tissues and organs, no development to the adult stage was reported, leaving open the question of whether a differentiated adult nucleus can be fully reprogrammed. Previously we reported the birth of live lambs after nuclear transfer from cultured embryonic cells that had been induced into quiescence. We suggested that inducing the donor cell to exit the growth phase causes changes in chromatin structure that facilitate reprogramming of gene expression and that development would be normal if nuclei are used from a variety of differentiated donor cells in similar regimes. Here we investigate whether normal development to term is possible when donor cells derived from fetal or adult tissue are induced to exit the growth cycle and enter the G0 phase of the cell cycle before nuclear transfer. Three new populations of cells were derived from (1) a day-9 embryo, (2) a day-26 fetus and (3) mammary gland of a 6-year-old ewe in the last trimester of pregnancy. Morphology of the embryo-derived cells (Fig. 1) is unlike both mouse embryonic stem (ES) cells and the embryo-derived cells used in our previous study. Nuclear transfer was carried out according to one of our established protocols1 and reconstructed embryos transferred into recipient ewes. Ultrasound scanning detected 21 single fetuses on day 50--60 after oestrus (
Table 1 ). On subsequent scanning at ~14-day intervals, fewer fetuses were observed, suggesting either mis-diagnosis or fetal loss. In total, 62% of fetuses were lost, a significantly greater proportion than the estimate of 6% after natural mating4. Increased prenatal loss has been reported after embryo manipulation or culture of unreconstructed embryos5. At about day 110 of pregnancy, four fetuses were dead, all from embryo-derived cells, and post-mortem analysis was possible after killing the ewes. Two fetuses had abnormal liver development, but no other abnormalities were detected and there was no evidence of infection.
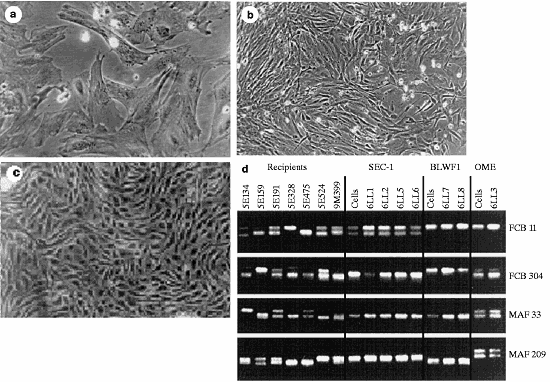
- Figure 1 Phase-contrast photomicrograph of donor-cell populations: a, Embryo-derived cells (SEC1); b, fetal fibroblasts (BLWF1); c, mammary-derived cells (OME). d, Microsatellite analysis of recipient ewes, nuclear donor cells and lambs using four polymorphic ovine markers22. The ewes are arranged from left to right in the same order as the lambs. Cell populations are embryo-derived (SEC1), fetal-derived (BLW1), and mammary-derived (OME), respectively. Lambs have the same genotype as the donor cells and differ from their recipient mothers.
Table 1 Development of embryos reconstructed with three different cell types Cell type No. of fused couplets (%)* No. recovered from oviduct (%) No. cultured No. of morula/blastocyst (%) No. of morula or blastocysts transferred? No. of pregnancies/ no. of recipients (%) No. of live lambs (%)? Mammary epithelium 277 (63.8)a 247 (89.2) -- 29 (11.7)a 29 1/13 (7.7) 1 (3.4%) Fetal fibroblast 172 (84.7)b 124 (86.7) --
2434 (27.4)b
13 (54.2)b34
64/10 (40.0)
1/6 (16.6)2 (5.9%)
1 (16.6%)°Embryo-derived 385 (82.8)b 231 (85.3) --
9290 (39.0)b
36 (39.0)b72
1514/27 (51.8)
1/5 (20.0)4 (5.6%)
0* As assessed 1 h after fusion by examination on a dissecting microscope. Superscripts a or b within a column indicate a significant difference between donor cell types in the efficiency of fusion ( P < 0.001 ) or the proportion of embryos that developed to morula or blastocyst (P < 0.001 ).? It was not practicable to transfer all morulae/blastocysts.
? As a proportion of morulae or blastocysts transferred. Not all recipients were perfectly synchronized.
° This lamb died within a few minutes of birth.
- Eight ewes gave birth to live lambs (Table 1, Fig. 2). All three cell populations were represented. One weak lamb, derived from the fetal fibroblasts, weighed 3.1 kg and died within a few minutes of birth, although post-mortem analysis failed to find any abnormality or infection. At 12.5%, perinatal loss was not dissimilar to that occurring in a large study of commercial sheep, when 8% of lambs died within 24 h of birth6. In all cases the lambs displayed the morphological characteristics of the breed used to derive the nucleus donors and not that of the oocyte donor (Table 2). This alone indicates that the lambs could not have been born after inadvertent mating of either the oocyte donor or recipient ewes. In addition, DNA microsatellite analysis of the cell populations and the lambs at four polymorphic loci confirmed that each lamb was derived from the cell population used as nuclear donor (
Fig. 1 ). Duration of gestation is determined by fetal genotype7, and in all cases gestation was longer than the breed mean (Table 2). By contrast, birth weight is influenced by both maternal and fetal genotype8. The birth weight of all lambs was within the range for single lambs born to Blackface ewes on our farm (up to 6.6 kg) and in most cases was within the range for the breed of the nuclear donor. There are no strict control observations for birth weight after embryo transfer between breeds, but the range in weight of lambs born to their own breed on our farms is 1.2--5.0 kg, 2--4.9 kg and 3--9 kg for the Finn Dorset, Welsh Mountain and Poll Dorset genotypes, respectively. The attainment of sexual maturity in the lambs is being monitored.
- Figure 2 Lamb number 6LL3 derived from the mammary gland of a Finn Dorset ewe with the Scottish Blackface ewe which was the recipient.
Table 2 Delivery of lambs developing from embryos derived by nuclear transfer from three different donor cells types, showing gestation length and birth weight Cell type Breed of lamb Lamb identity Duration of pregnancy (days)* Birth weight (kg) Mammary epithelium Finn Dorset 6LL3 148 6.6 Fetal fibroblast Black Welsh
Black Welsh
Black Welsh6LL7
6LL8
6LL9**152
149
1565.6
2.8
3.1Embryo- derived Poll Dorset
Poll Dorset
Poll Dorset
Poll Dorset6LL1
6LL2***
6LL5
6LL6?149
152
148
1526.5
6.2
4.2
5.3* Breed averages are 143, 147 and 145 days, respectively for the three genotypes Finn Dorset, Black Welsh Mountain and Poll Dorset. ** This lamb died within a few minutes of birth.
*** These lambs were delivered by caesarian section. Overall the nature of the assistance provided by the veterinary surgeon was similar to that expected in a commercial flock.
- Development of embryos produced by nuclear transfer depends upon the maintenance of normal ploidy and creating the conditions for developmental regulation of gene expression. These responses are both influenced by the cell-cycle stage of donor and recipient cells and the interaction between them (reviewed in ref. 9). A comparison of development of mouse and cattle embryos produced by nuclear transfer to oocytes10,11 or enucleated zygotes12,13 suggests that a greater proportion develop if the recipient is an oocyte. This may be because factors that bring about reprogramming of gene expression in a transferred nucleus are required for early development and are taken up by the pronuclei during development of the zygote.
- If the recipient cytoplasm is prepared by enucleation of an oocyte at metaphase II, it is only possible to avoid chromosomal damage and maintain normal ploidy by transfer of diploid nuclei14,15, but further experiments are required to define the optimum cell-cycle stage. Our studies with cultured cells suggest that there is an advantage if cells are quiescent (ref. 1, and this work). In earlier studies, donor cells were embryonic blastomeres that had not been induced into quiescence. Comparisons of the phases of the growth cycle showed that development was greater if donor cells were in mitosis16 or in the G1 (ref. 10) phase of the cycle, rather than in S or G2 phases. Increased development using donor cells in G0, G1 or mitosis may reflect greater access for reprogramming factors present in the oocyte cycoplasm, but a direct comparison of these phases in the same cell population is required for a clearer understanding of the underlying mechanisms.
- Together these results indicate that nuclei from a wide range of cell types should prove to be totipotent after enhancing opportunities for reprogramming by using appropriate combinations of these cell-cycle stages. In turn, the dissemination of the genetic improvement obtained within elite selection herds will be enhanced by limited replication of animals with proven performance by nuclear transfer from cells derived from adult animals. In addition, gene targeting in livestock should now be feasible by nuclear transfer from modified cell populations and will offer new opportunities in biotechnology. The techniques described also offer an opportunity to study the possible persistence and impact of epigenetic changes, such as imprinting and telomere shortening, which are known to occur in somatic cells during development and senescence, respectively.
- The lamb born after nuclear transfer from a mammary gland cell is, to our knowledge, the first mammal to develop from a cell derived from an adult tissue. The phenotype of the donor cell is unknown. The primary culture contains mainly mammary epithelial (over 90%) as well as other differentiated cell types, including myoepithelial cells and fibroblasts. We cannot exclude the possibility that there is a small proportion of relatively undifferentiated stem cells able to support regeneration of the mammary gland during pregnancy. Birth of the lamb shows that during the development of that mammary cell there was no irreversible modification of genetic information required for development to term. This is consistent with the generally accepted view that mammalian differentiation is almost all achieved by systematic, sequential changes in gene expression brought about by interactions between the nucleus and the changing cytoplasmic environment17.
- Methods
- Embryo-derived cells were obtained from embryonic disc of a day-9 embryo from a Poll Dorset ewe cultured as described1, with the following modifications. Stem-cell medium was supplemented with bovine DIA/LIF. After 8 days, the explanted disc was disaggregated by enzymatic digestion and cells replated onto fresh feeders. After a further 7 days, a single colony of large flattened cells was isolated and grown further in the absence of feeder cells. At passage 8, the modal chromosome number was 54. These cells were used as nuclear donors at passages 7--9. Fetal-derived cells were obtained from an eviscerated Black Welsh Mountain fetus recovered at autopsy on day 26 of pregnancy. The head was removed before tissues were cut into small pieces and the cells dispersed by exposure to trypsin. Culture was in BHK 21 (Glasgow MEM; Gibco Life Sciences) supplemented with
L -glutamine (2 mM), sodium pyruvate (1 mM) and 10% fetal calf serum. At 90% confluency, the cells were passaged with a 1 : 2 division. At passage 4, these fibroblast-like cells (Fig. 1) had modal chromosome number of 54. Fetal cells were used as nuclear donors at passages 4--6. Cells from mammary gland were obtained from a 6-year-old Finn Dorset ewe in the last trimester of pregnancy18. At passages 3 and 6, the modal chromosome number was 54 and these cells were used as nuclear donors at passage numbers 3--6.- Nuclear transfer was done according to a previous protocol1. Oocytes were recovered from Scottish Blackface ewes between 28 and 33 h after injection of gonadotropin-releasing hormone (GnRH), and enucleated as soon as possible. They were recovered in calcium- and magnesium-free PBS containing 1% FCS and transferred to calcium-free M2 medium19 containing 10% FCS at 37 °C. Quiescent, diploid donor cells were produced by reducing the concentration of serum in the medium from 10 to 0.5% for 5 days, causing the cells to exit the growth cycle and arrest in G0. Confirmation that cells had left the cycle was obtained by staining with antiPCNA/cyclin antibody (Immuno Concepts), revealed by a second antibody conjugated with rhodamine (Dakopatts).
- Fusion of the donor cell to the enucleated oocyte and activation of the oocyte were induced by the same electrical pulses, between 34 and 36 h after GnRH injection to donor ewes. The majority of reconstructed embryos were cultured in ligated oviducts of sheep as before, but some embryos produced by transfer from embryo-derived cells or fetal fibroblasts were cultured in a chemically defined medium20. Most embryos that developed to morula or blastocyst after 6 days of culture were transferred to recipients and allowed to develop to term (Table 1). One, two or three embryos were transferred to each ewe depending upon the availability of embryos. The effect of cell type upon fusion and development to morula or blastocyst was analysed using the marginal model of Breslow and Clayton21. No comparison was possible of development to term as it was not practicable to transfer all embryos developing to a suitable stage for transfer. When too many embryos were available, those having better morphology were selected.
- Ultrasound scan was used for pregnancy diagnosis at around day 60 after oestrus and to monitor fetal development thereafter at 2-week intervals. Pregnant recipient ewes were monitored for nutritional status, body condition and signs of EAE, Q fever, border disease, louping ill and toxoplasmosis. As lambing approached, they were under constant observation and a veterinary surgeon called at the onset of parturition. Microsatellite analysis was carried out on DNA from the lambs and recipient ewes using four polymorphic ovine markers22.
- Correspondence should be addressed to I.W. (e-mail Ian.Wilmut@bbsrc.ac.uk).
- We thank A. Colman for his involvement throughout this experiment and for guidance during the preparation of this manuscript; C. Wilde for mammary-derived cells; M. Ritchie, J. Bracken, M. Malcolm-Smith, W. A. Ritchie, P. Ferrier and K. Mycock for technical assistance; D. Waddington for statistical analysis; and H. Bowran and his colleagues for care of the animals. This research was supported in part by the Ministry of Agriculture, Fisheries and Food. The experiments were conducted under the Animals (Scientific Procedures) Act 1986 and with the approval of the Roslin Institute Animal Welfare and Experiments Committee.
- Campbell, K. H. S., McWhir, J., Ritchie, W. A. & Wilmut, I. Nature 380, 64--66 (1996).
- Solter, D. Nature 380, 24--25 (1996).
- Gurdon, J. B., Laskey, R. A. & Reeves, O. R. J. Embryol. Exp. Morph. 34, 93--112 (1975).
- Quinlivan, T. D., Martin, C. A., Taylor, W. B. & Cairney, I. M. J. Reprod. Fert. 11, 379--390 (1966).
- Walker, S. K., Heard, T. M. & Seamark, R. F. Therio 37, 111--126 (1992).
- Nash, M. L., Hungerford, L. L., Nash, T. G. & Zinn, G. M. Vet. Rec. 139, 64--67 (1996).
- Bradford, G. E., Hart, R., Quirke, J. F. & Land, R. B. J. Reprod. Fert. 30, 459--463 (1972).
- Walton, A. & Hammond, J. Proc. R. Soc. B 125, 311--335 (1938).
- Campbell, K. H. S., Loi, P., Otaegui, P. J. & Wilmut, I. Rev. Reprod. 1, 40--46 (1996).
- Cheong, H.-T., Takahashi, Y. & Kanagawa, H. Biol. Reprod. 48, 958--963 (1993).
- Prather, R. S. Biol. Reprod. 37, 859--866 (1987).
- McGrath, J. & Solter, D. Science 226, 1317--1318 (1984).
- Robl, J. M. J. Anim. Sci. 64, 642--647 (1987).
- Campbell, K. H. S., Ritchie, W. A. & Wilmut, I. Biol. Reprod. 49, 933--942 (1993).
- Barnes, F. L. Mol. Reprod. Dev. 36, 33--41 (1993).
- Kwon, O. Y. & Kono, T. J. Reprod. Fert. Abst. Ser. 17, 30 (1996).
- Gurdon, J. B. , Oxford University Press, Oxford 1974.
- Finch, L. M. B. Biochem. Soc. Trans. 24, 369S (1996).
- Whitten, W. K. & Biggers, J. D. J. Reprod. Fertil. 17, 399--401 (1968).
- Gardner, D. K., Lane, M., Spitzer, A. & Batt, P. A. Biol. Reprod. 50, 390--400 (1994).
- Breslow, N. E. & Clayton, D. G. J. Am. Stat. Assoc. 88, 9--25 (1993).
- Buchanan, F. C., Littlejohn, R. P., Galloway, S. M. & Crawford, A. L. Mammal. Gen. 4, 258--264 (1993).
Nature © Macmillan Publishers Ltd. 1997
Registered No. 785998 England.